��ɫư����ձ�����ʳ���ܵij����о�
�ձ������Asterococcus muratae Kuwana�����ٺ����ٺ��ó����ֲ�������ڶ࣬�ҹ��ѱ����ļ���ֲ�����17��38��[1]�����кɻ�����Magnolia grandiflora L.���¼�Rosa chinensis Jacq.������Cinnamomum camphora L.�����ù�Cinnamomum japonicum Sieb.��������Acer buergerianum Miq.����Koelreuteria paniculata Laxm.������Pterocarya stenoptera C. DC.��СҶŮ��Ligustrum quihoui Carri��re���ձ�ɺ����Viburnum awabuki K. Koch��ɽ��Camellia japonica L.��Ϊ�������ֹ���ֲ��ó��Գɳ��������ʳ����ֲ������ɡ�֦��֭Һ��ͬʱ��й��¶��Honeydew����������������е�����Ҫ��������[2][3]���ɻ�����[4]�����ù��[5]����ú�۲�����Ҫԭ�����ؽ������־��۵Ĺ��ͼ�ֵ��
�ó����������һ�귢��һ��������3��4��Խ���ѷ���Ϊ���棬1��������֦Ҷ������ȡʳ��1��2 d���γɼ�Ӳ��ǹ̶���֦��������[6]������Ҫ��ȷ�����ձ������ķ������ڣ�����δ�γɽ��֮ǰ����ʩҩ���ء��ֽ���һ�߹ܻ���λ�����ձ���������Ҫ�ֶ���ͨ���ؼ�֦������ʩҩ��ȥ��ú�ۺڰ߽��з��Σ�һ���ؼ�֦��������Ӱ���־���Ч��������ȥ��ú�ۺڰ߲��ܷ�ֹ�����ձ���������ľ��Ӱ�졣�Ҹó��γɽ�Ǻ�һ��ҩ�����Դ���ʵ�ʲ���������Ϊ�ﵽ����Ч����ʩҩʱ�����üװ��ס���Ч�ס������ֹ��ȸ߶����߲�������ҩ��[4]����������̬����˲���Ӱ�졣Ϊ�ƽ������д���������̬�ֳ��й����������о�������������������ձ�����
�������أ��ձ�������ư��������к�Ե��ư��Chilocorus rubidus Hope��������ư��Menochilus sexmaculata Fabricius��������㴽ư��Chilocorus hupehanus Miyatake������ư��Propylea japonica Thunberg[7]����ɫư��Harmonia axyridis Pallas���컷ư��Rodolia limbata Motschulsky[8]�ȡ��й�ư������жԽ�dz�IJ�ʳ���ܵ��о����н�Ϊ�꾡�ı��������Ե��ư��ɳ��1��ƻ�����Rhodococcus sariuoni Borchsenius����������ʳ��Ϊ103.0ͷ[9]������ư��ɳ��������Kuwanaspis vermiformis Takahashi 1��2�������Լ��Ƴɳ�������ʳ���ֱ�Ϊ221.3��127.8��51.6ͷ[10]��ϸԵ��ư��Chilocorus circumdatus Gyllenhal�ɳ��ɣ���Pseudaulacaspis pentagona Targioni�Ƴɳ桢�Ѽ����������վ����ʳ����ֵ�ֱ�Ϊ11.07ͷ��140.91����65.00ͷ[11]����ư������ж��ձ������Ŀ�������������ȷ��������Ҳ��һ���о�������������ֶη����ձ���������ر���������ɫư�������Ӧ�ԡ���ֳ��ǿ����Ʒ���̶ȸߵ��ص㣬����2021��1��2022��4����ʵ�����ڹ۲�����ɫư��ɳ���ձ������1������IJ�ʳ���ã������о��䲶ʳ���ܷ�Ӧ��������Ϊ�ƶ�������δ�ʩ�����ձ�������ṩ�������ݣ��Ӷ���߳����־���Ʒ�ʡ�
1�������뷽��
1.1 �������
������ɫư��ɳ����������з羰�ֿ�ѧ�о�Ժ��з���ʵ�������ڷ���Ʒϵ��ʹ�������ڲ϶���Vicia faba L.�ϵ��㶹��Acyrthosiphon pisum Harris�����������ձ�������ѿ�ɼ�����������������ֲ���е��������������������ù��ϣ������£�25��1�棩��������16L:8D������������ѡȡ�����еij���1������������顣����������£�25��1�棩��������16L:8D�½��С�
1.2 ��ɫư��Բ�ͬ�ܶ��ձ������Ĺ��ܷ�Ӧ
������������ֱ��90 mm����15 mm������1Ƭֱ��Ϊ60 mm����ֽ��ʹ�����ܵ�3�����ˮ���б�ʪ�����������߱������ձ������1�����棬�����ձ�������ܶ��ݶȷֱ�Ϊ30��70��110��150��190��230ͷ/����6���ݶȣ�ÿ�����ظ�5�Ρ�������24 h�����ɫư��ɳ�����������ڣ�ÿ�����������1ͷ�������parafilm���Ĥ�ܷ�����������������С�ף���֤�������ڵ�ͨ���ԡ�24 h��۲첢��¼ÿ���ձ������ʣ�������
1.3 ��ɫư��������ܶȵĹ��ܷ�Ӧ
ÿ���������з���1Ƭֱ��Ϊ60 mm����ֽ��ʹ�����ܵ�3��ˮ����ֽ�Ͻ��б�ʪ��������110ͷ�ձ������1�����档������24 h����ɫư��ɳ�ֱ�����1��2��3��4��5ͷ/��5��������ÿ�����ظ�5�Ρ�24 h��۲첢��¼ÿ���ձ������ʣ�������
1.4 ����ͳ�������
1.4.1 ��ɫư��Բ�ͬ�ܶ��ձ������Ĺ��ܷ�Ӧ
��ɫư��ɳ�Բ�ͬ�ܶ��ձ������1������IJ�ʳ����ϵ����Holling-��Բ�̷���[12]false������ϣ�ʽ��NaΪ����ʳ������������aΪ��ʳ�߶������˲ʱ�����ʣ�NΪ�����ܶȣ�ThΪ��ʳ1ͷ��������ʱ�䣬TΪ��ʳ�߲�ʳ�������ʱ�䣨ȡT=1 d��24 h������������ղ�ʳ����ʾΪ1/Th����ʳЧ��Ϊa/Th[13]��
1.4.2 ��ɫư����ձ�������Ѱ��ЧӦ
��ɫư����ձ�������Ѱ��ЧӦ���÷���false��ʽ��SΪѰ��ЧӦ���������ͬ�ϡ�
1.4.3 ��ɫư��������ܶȵĹ��ܷ�Ӧ
��ɫư��������ܶȵĹ��ܷ�Ӧ���ò�ʳ������E���ۣ��������ݲ���Hassell-Varleyģ��[14]������ϣ�false������false��EΪ��ʳ�����ʣ�QΪ��Ѱ������PΪ��ʳ���ܶȣ�mΪ���ų������������ͬ�ϡ�
1.4.4��ɫư�����Ѱ���ܶȹ���
��ʳ���������IJ�ʳ��Ϊֻ������������ܶ������²��ܷ������Ч�ܡ�����Na=a∙exp��b∙N-1��������ɫư������Ѱ���ܶȡ�ʽ��aΪ������ʳ����bΪ����״̬�µ����Ѱ���ܶȣ��������ͬ�ϡ��������ݾ���Excel��¼�봦����������SPSS��Ϲ��ܷ�Ӧ���̡�
2��ʵ����
2.1 ��ɫư��Բ�ͬ�ܶ��ձ������Ĺ��ܷ�Ӧ
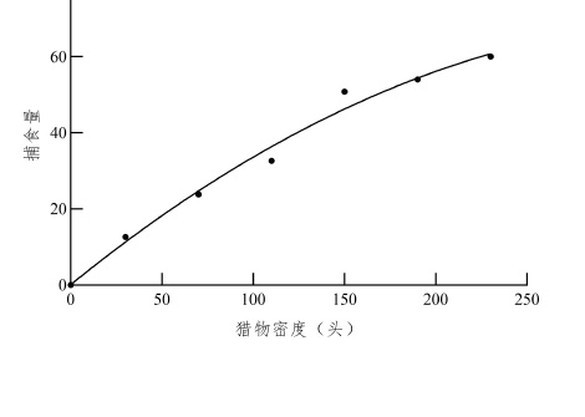
��ɫư��IJ�ʳ�����ձ�������ܶ����Ӷ��������ձ�������ܶȳ�������ʱ����ɫư��IJ�ʳ������ƽ������ͼ1����

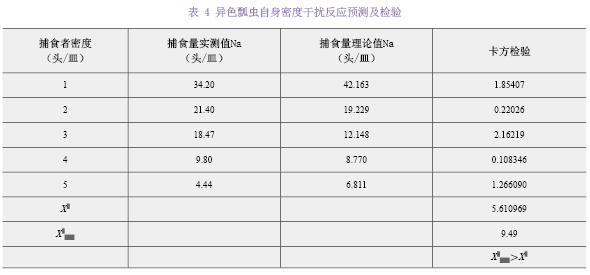
��Holling-�����Ի������1/Na��y����1/N��x���������Թ�ϵ��y=2.1773x+0.008����ò�ʳ���ܷ�Ӧ����Ϊfalse����R2=0.9903����˲ʱ������a=0.4593����ʳ�ߴ���1ͷ���������ѵ�ʱ��Th=0.008 d��a/Th=57.3954��˵����ɫư����ձ���������������ǿ��1ͷ��ɫư����24 h�ڵ��������ʳ��Na max=1/Th=124.9653ͷ������1��
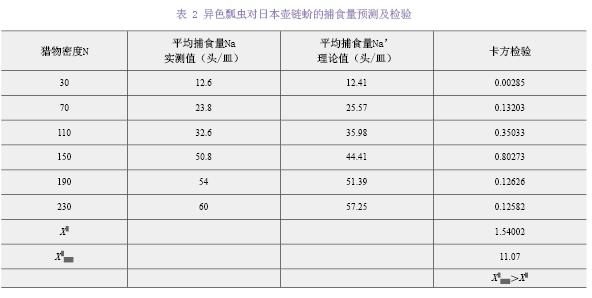
����ɫư��ƽ����ʳ��������ֵ��ʵ��ֵ������2�����п������飬���X2=1.54002��X2��5��0.05��=11.07��ˮƽ�ϲ����������ԣ����ܾܾ�ԭ���裬��Holling-��ģ���ܽϺõط�Ӧ��ɫư��ɳ���ձ������IJ�ʳ���ܷ�Ӧ��
2.2 ��ɫư����ձ�������Ѱ��ЧӦ
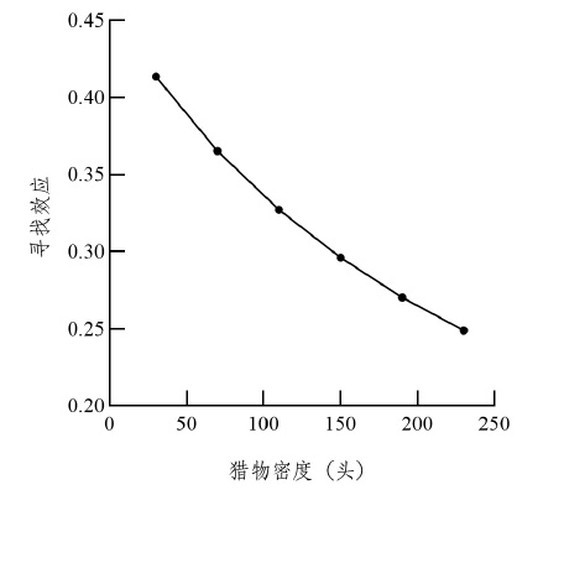
��ɫư����ձ�������Ѱ��ЧӦ����Ϊfalse�������ʾ�������ձ��������ܶ���30ͷ/�����ӵ�230ͷ/����ɫư����ձ�������Ѱ��ЧӦ��0.4137����Ϊ0.2489��ͼ2����
2.3 ��ɫư��������ܶȵĹ��ܷ�Ӧ
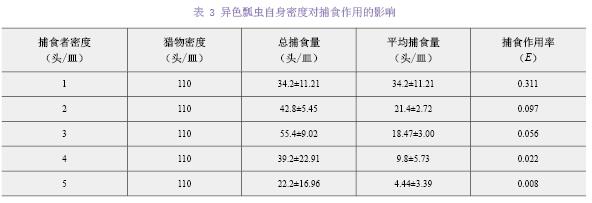
���ռ�������ܶȣ��ձ������110ͷ��һ������ɫư����ձ�������ƽ����ʳ�����������ܶȵ��������٣���ʳ�����ʣ�E��Ҳ��Ӧ���ͣ���3����
��ɫư�������ܶȸ������ÿ���Hassell-Varley�ĸ��ŷ�Ӧģ�ͽ�����ϣ��ó���������Q=0.38329������ϵ��m=2.1327�������ܶȸ��ŷ���Ϊ��false��r=0.9609��
����1ͷ��ɫư�沶ʳ��ʵ��ֵ������ֵ�Ŀ������飬X20.05��X2����ʵ��ֵ������ֵ�����Բ��죬��Hassell-Varley�ĸ��ŷ�Ӧģ���ܺܺõط�ӳ��ɫư�沶ʳ�ձ����������е������ܶȸ������������4��
2.4 ��ɫư�����Ѱ���ܶȹ���
��1ͷ��ɫư��ɳ��ڲ�ͬ�ܶȵ��ձ������1�������²�ʳ��ʵ�����ݹ������Ѱ���ܶȣ����Na=379.79∙exp��-52.38N-1��������������ʳ��a=379.79������״̬�����Ѱ���ܶ�Ϊb=52.38�����ⶨ��ɫư��ɳ���ձ������1��������Ƶ��溦�ȿ���Ϊ1��52������ʵ��������ȶ���յĿռ��ڽ��У��������·�̵�ʵ���е�����ֲ�������ɵĸ��ӻ���ʵ�����������룬�ֿ��ǵ������鷽����Ϊ�����ձ������Ⱥ��Σ�����ص㣬��Ϊ�˽���ܽϺô�����ɫư����ձ������1������IJ�ʳ��Ϊ��
3�� ��������
�ֽ��о���ʳ���������Ժ���IJ�ʳ������Ҫͨ���о��䲶ʳ���ܷ�Ӧ���У��ܹ���Ч������������Ŀ�꺦��Ŀ���Ч�����ڱ������У���ɫư�沶ʳ�ձ��������������ձ�������ܶ�֮�����Holling-���ܷ�Ӧģ�ͣ�����ʳ�����ձ�������ܶ�������������ձ�������ܶȴﵽһ���̶Ⱥ���ɫư�沶ʳ���������ӡ��ɴ˿ɼ������̵���̬�����е��ձ������ڳ�������ʱ����������ɫư��ɳ���ձ������Ͷ�ű���1��52������������Ͷ�š�
������Ϊʹ��������η��������ձ�������ṩ����˼·������һ���̶��ϼ��������ũҩ��ʹ�öԻ�����ɵĸ�������ʵ�������µIJ�ʳ��������Ȼ�����д����Žϴ��࣬����Ӧ��һ���о���ɫư�治ͬ���ںͲ�ͬ�¶������¶��䲶ʳ�ձ�������Ӱ�죬���õ�ָ������������η�������ú�۲���
��������Ŀ�������г��й���������Ŀ����ú�۲���Ҫ�β����桪���ձ���������������о������á����ǹܿ���2020�ڣ�08���ţ���
�ο����ף�
[1] �����,ë��Ԫ,��ƽ.�ձ������������μ����о���չ[J].����ũҵ��ѧ,2013,41(04):1521-1523+1525.
[2] ��һ��,������,��־ƽ,Ѧ��,����.����ҩ����������ú�۲���ҩЧ����[J].�й�ɭ�ֲ���,2014,33(03):34-37.
[3] ������,��־��.����������ú�۲����ձ�������Σ���������[J].������ҵ�Ƽ�,2017,44(05):32-34.
[4] ����,����,���н�,����.���ɴ��עҩ����Σ�����������ձ������[J].�й�ɭ�ֲ���,2006(02):34-37.
[5] ����,������,������.����������ù�ú�۲����ձ��������Ĺ�ϵ�о�[J].ֲ��ҽ��,2021,34(06):24-27.
[6] �����.�ٺ��������ϰ�Լ������о�����[J].ֲ�ﱣ��,1985(03):17-19.
[7] ����Ȫ,��ѵ��,����,¬����,��ά.���Ӳ��溦����ij�������[J].ֲ�ﱣ��,2011,37(01):132-134+137.
[8] �ν�ѫ,����,����.�ٺ����ķ������ɼ������[J].����֪ʶ,1994(01):22-24.
[9] ����,����,���Ӿ�,�����.��Ե��ư���ƻ����ʳ���ܵij����о�[J].�й�ֲ������,2022,42(11):18-21.
[10] Ҷ�帣.����ư��ɳ��������IJ�ʳ���ܷ�Ӧ[J].�ӱ���ҵ�Ƽ�,2012(04):1-3.
[11] ����÷,�Ʒ�.ϸԵ��ư�沶ʳɣ���ܷ�Ӧ��Ѱ��ЧӦ[J].�й�ֲ������,2014,34(08):18-22.
[12] Holling C S. The functional response of invertebrate predators to prey density[J]. The Memoirs of the Entomological Society of Canada, 1966, 98(S48): 5-86.
[13] ����,������,����ǿ,���,����.��ϣ�������й��濦ľʭ�ɳ�IJ�ʳ����[J].ֲ�ﱣ��,2012,38(05):112-114.
[14] Hassell M P, Comins H N. Sigmoid functional responses and population stability[J]. Theoretical Population Biology, 1978, 14(1): 62-67.
��Ȩ��ԭ�������С�����Ϊ������Ȩ������ϵ����ɾ����


��Ա֮��

- ���죺�����з羰��ѧ���а죺������������ѯ����˾
- ѧ��绰��(023)68801472 63673736 63673738 63892447QQ:944865067
- ��ַ�������Ԭ·205�����ͻ���A��3¥
- ��ICP��11002947��-1 �幫������ 50010302002828��